2007/01/29 Monday
2007/01/29(Mon)
��
 ������ij��������ܤؤ����������Ϥ�Ļ�ˤ���ΤȤ��Ƥ⡢�Ǥϡ��ɤ����ơ��Ϥ�Ļ���������ʤ��ܼ���ˤޤǡ����äƤ��Ƥ��ޤä��Τ����ˤĤ��Ƥϡ��ʤ����ĤäƤ���ΤǤ���������Ȥ���Ǥϡ��ϥ�������Ū�����Ƥ���ΤǤϤʤ����ȡ���������⤢��褦�Ǥ���
������ij��������ܤؤ����������Ϥ�Ļ�ˤ���ΤȤ��Ƥ⡢�Ǥϡ��ɤ����ơ��Ϥ�Ļ���������ʤ��ܼ���ˤޤǡ����äƤ��Ƥ��ޤä��Τ����ˤĤ��Ƥϡ��ʤ����ĤäƤ���ΤǤ���������Ȥ���Ǥϡ��ϥ�������Ū�����Ƥ���ΤǤϤʤ����ȡ���������⤢��褦�Ǥ���
�����ȡ�2004ǯ���¸���Ļ����ե륨����ή���ϤǺν����줿�����Х���blow fly���फ���H5N1��������ե륨�����륹�θ��Ф�ʬΥ�פϡ���Ω�����ɸ����θ���Ǥ����������ˤ����Ƥϡ���ʬ�����Į�ȵ�����ð��Į���ߵ��˳�ư����ϥ���κν���Ԥ��������ν����Τ���H5N1��������ե륨�����륹�θ��Ф�ʬΥ���ߤ��Ȥ��������������Х������֥������Х��ΰ����礫�顢H5N1(A/blow fly/Kyoto/93/2004)�������Ф��줿�Ȥ��Ƥ��ޤ���
�ޤ�����ǯ9��ˡ�����ɥͥ��������祯���㥫�륿�Υ���ޥ����(Universitas Gadjah Mada)�ǡ�Ʊ�ͤˡ��ϥ���Ļ�����륹�����夷�Ƥ����Τ�ȯ�������Ȥ���ƻ�⤢�ä��褦�Ǥ���
����ϡ�����ޥ���ؤ�Wasito��θ���ˤ���Τǡ������Ǥϡ��ϥ��ξò��ﴱ��Ƶ۴ﴱ���顢�����륹�Ф����ȤΤ��ȤǤ���(�����������ΤȤ��Υ����륹�ϡ��ޤ�����The Influenza Sequence Database�פˤϡ���Ͽ����Ƥ��ʤ��褦�Ǥ��͡�
����㥫�륿�ݥ��Ȼ�˸�ä������Ǥϡ��ظ���ǭ�⡢���뤤�ϡ��ϥ��Ǥ����⡢Ļ����ե륨�����륹�˴��������ǽ�������롣�١ؤ����θ���Ǥϡ��ϥ��ϡ�H5N1�����륹��mechanical transmitter(��ã����)���ꤦ�뤳�Ȥ����Ƥ��롣��( Our results suggest it is possible that blow flies could become a mechanical transmitter of H5N1 influenza virus. )�ȸ�äƤ���褦�Ǥ���)
���ȡ�Scientist warns of bird flu in flies ��
��Cats and Dogs and Flies, Oh My! ��
�Ǥϡ�������ɥ��쥹(������ɥ쥹)�ǤϤɤ����Ȥ����ȡ�����ˤĤ��Ƥ⡢��������Υ֥���������������ɥ��쥹�ܼˤ��Ф����ɱ־�����̰����ϡ�����٤������Ǥ�Ҥ٤��褦�ˡ������ܼˤ����������ܼˤ���١������ꥹ�����㤤�ȤϤ������۵���ʬ�Υ�å���ϡ��ϥ��ο������ɤ��ۤɡ��٤����Ϥʤ����Ȥ������ȤΤ褦�Ǥ���
����ˤϡ�������ɥ쥹�δ��������ˤϡ��۰����������������ʤɡ����������ʼ���Ϥ����ΤΡ�������ɥ쥹�ܼˤ��������Ū�ϡ������Ϥ��ɴ��ȸ��������Ǥ��ꡢ��Ȥ���ɱ�������Ū�ˤϵ�ǽ���Ƥ��ʤ��Ȥ����ո��⤢�뤯�餤�Ǥ���
��Avian Influenza (Bird Flu): Agricultural and Wildlife Considerations �פǤϡ������β�ǽ���Ȥ��ơ������װ���Ƥ���褦�Ǥ���
��.ĻƱ�Τ����ܤ��Ƥ�����ϡ����������⤢�ꤦ�롣
2.�Ƶ۴�ʬ��ʪ��ʵ�ʤɤα�ʪ�ؤ�ľ��Ū�ܿ���
3.��ľ�����ϡ������äƤ��ʤ���
4.�ܤν�����Ǥϡ����δ������Ǥ����롣
����١�����������줿��硢���ؤδ��������롣
���������ܤ�����������ư���뤳�Ȥˤ�봶�����硣
���������䡢���Ȱ��ΰ��ࡦ���ʤɴ�������Ȥʤ��ΤΡ��������ư
������Ļ���Ļ�ؤ��ܿ�
���߿��ʵ�ˤ�����
�ܼ���Υϥ�
���Τ������Ǹ�Ρַܼ���Υϥ��פˤĤ��Ƥϡ���Garbage flies (suspected of transmitting the virus during the 1983-1984 epidemic in Pennsylvania) �פȤ��ꡢ��1983ǯ����1984ǯ�ˤ����ƤΡ��ڥ�٥˥����ǤΣ�5��1(A/Gull/Pennsylvania/4175/83)��������θ����ȿ�¬����롣�פȤ���ޤ���
���Υڥ�٥˥��Ǥλ���ˤĤ��ơ����Υ����ȡ�Wishful Thinking�פǤϡ���Ļ����ե륨�����륹�α������줿�Ӥο夬���ϲ�������߽Ф��Ƥ������פȤ��Ƥ��ޤ���������Ǥϡ���䡢��¬������褦�ˤ⸫���ޤ���
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/25 Thursday
2007/01/25(Thu)
��
 �������ij��Ф�H5N1���ϡ��ɤΤ褦�ʥ������ɤäơ����ܤˤ��ɤ��夤����?��
�������ij��Ф�H5N1���ϡ��ɤΤ褦�ʥ������ɤäơ����ܤˤ��ɤ��夤����?��
�Ȥ����褦�ʡ����䤤��碌����¿������ޤ��Τǡ��狼��䤹���Ͽޤ��˼����Ƥ����ޤ���
���Ͽޤϡ������Ǥ�����
��H5N1 Wild Bird Flu Outbreaks��February 2006�פ�����
�ϿޤΥޡ����ˤĤ���
�Ŀ�2005/05��������2005/07���Ͳ���2005/07���п�2005/08/15���ֿ�2005/08/31�ͻ翧2005/09����2005/10�Ͷ���2005/11�ͳ���2005/12�ͳ쿧2006/01��2006/02���ӿ�
�ޤ����Ͽޤο�������Ĵݤ��ij��Фξ��ǡ����줬����ü�����٥ꥢ������̾夷�Ƥ��顢����ʬ����������
�������ϡ��Х륫��Ⱦ���Фơ��衼���åѤ��Ϥꡢ�������餵�������ƥ��եꥫ�������ץȤ��Ϥ��Τȡ�
���⤦�����ϡ������������ڹ�����ܤؤȡ�����륳������
������ޤ���
 �������Ͽޤ������Ͽޤǡ��濴���֤��ϰ褬���ij��Фǡ��������顢���礷�Ƥ��ä��Ȥ����櫓�Ǥ��͡�
�������Ͽޤ������Ͽޤǡ��濴���֤��ϰ褬���ij��Фǡ��������顢���礷�Ƥ��ä��Ȥ����櫓�Ǥ��͡�
2005ǯ5��4�����ij��ФǤ�H5N1ȯ�����κ��𤷤����ͤ������2005ǯ5��23���Υ֥����������ҥȴ�����Ļ����ե륨�ϡ��ܳ�Ū������ή�Ԥ��ʳ��������פ˽�Ƥ��ޤ��Τǡ������Ȥ���������
�ޤ������λ����ǡ����Υ֥���������ͽ¬�������Ȥ������θ塢���٤������äƤ��ޤä��Ȥ����Τϡ������ʷ�̤Ǥ�����
���������ǯ�����̤��¤٤ޤ��ȡ������ΤȤ���Ǥ���
�ʤ��������Ǥδ����ˤĤ��Ƥϡ��������ij��������Ǥʤ���ʡ�����⡢�ޤߤޤ���
2005/05-����ij��� �仡�Ѣ�
2005/06-����� �����ҿ��
2005/08-��롢�����ե����������������鶼���ҿ �ǻ��Ԣ�
2005/10-�ȥ륳���롼�ޥ˥�������������������ɡ����內�ظż���ҿ ���¹��ûԡ��µ��� ŷĹ�ԡ������ ��߬�Ѣ�
2005/11-�����ǫ�ʡ������ʡ����̾ʡ����Űݸ㼤(��������)����ҿ �����̾� ��ǫ�Ʋ�²����ҿ������ʢ�
2005/12-�����饤�ʡ�������ʡ������ʡ�ʡ���ʢ�
2006/01-���饯�������ʡ����Ԣ�
2006/02-���ꥷ�㡢�����ꥢ���ե���ɥ��ġ��������ȥꥢ�����른�����ʥ������ꥢ���˥������롢�����ץȡ����������ϥ���֥륬�ꥢ�������٥˥��������Х������ܥ��˥����إ�ĥ����ӥʡ�����ѥ����������(����)���ʢ�
2006/03-���ե��˥��������饨�롢�ѥ쥹���ʡ���������롼����Х˥���������Х��������ӥ������ƥͥ������ݡ����ɡ��ǥ�ޡ������������������ʡ��峤��
2006/04-�֥륭�ʥե����������ȥ��ܥ����롢���ץ��������ꥹ��
2006/06�����圳�ʿ�����ˢ�
2006/07-���ڥ����
2006/10-���������
2006/11-�ڹ��
2007/01-���ܢ�?
���Τۤ��ˡ����ͻ����Ȥ��Ƥϡ�
�����Ļ����ե륨��ȯ��������ȯ����Į¼̾������
�䡢
�����ȡ�������Ļ����ե륨����ʬ�ۿ���
��
����ή���϶�ʬ�ۼ��տ���
��
�����ʹ�����ή���־�ʬ�ۿ���
�⤴����
�ʾ�Τ褦�ˡ��ij���������������ä����Ȥϡ��狼��ޤ����������⤽�⡢�����ij��������ɤ������褿�Τ����ˤĤ��Ƥϡ��������ʤ�����ޤ�Ƥ���褦�Ǥ��͡�
�鷺���ˡ���Probable H5N1 in Sicily Italy�פʤɡ�Recombinomics�ε����ˤ����ơ������ij����Υ��ꥸ��ϡ�����ɤˤ���Ȥ��Ƥ���ΤǤ��������Υ���ɤˤ����ơ��ο��Υ����륹�θ��Ф��Ǥ��Ƥ��ʤ��褦�Ǥ��͡�
���ʤߤˡ���Bird Flu in Migrating Bar Headed Geese from India�פǤϡ���������ǯ���������ij��Ф�ȯ�����줿Ļ����ե륨�ϡ��������ʿ��������Ϥ�Ļ�ˤ���ΤȤθ����Ƥ��ꡢ2005ǯ����ˡ�����ɤ� Uttar Pradesh�ϰ�Ǥ��ڡ��ҥȡ������δ����䡢�˥塼�ǥ�ǤΥҥȤο����(ή����Ǿ�Կ����)��ή�Ԥȴ�Ϣ������Ȥ��Ƥ��ޤ���
�ޤ�����������ǯ�ˤϡ�����ɤǡ��ȶ٤�ȶٽ����Ԥ��齸������������Ǥ��ä����Ȥ⡢�ط����Ƥ���ΤǤϤʤ����ȤߤƤ��ޤ���
����ɤǤϡ���������ǯ�ˡ����ͤβȶٽ����Ԥη������顢H5N1���Τ����Ĥ��ä����ȤǶ�ˤʤä�ǧ��Ƥ��뤫��Ǥ���
�����οͤϡ���������ε����ǡ����Ǥ��������ΤȤΤ��ȤǤ���
�������������ο����ȤƤ⡢�ξڤ�����櫓�ǤϤʤ����ޤ��ޤ����ʤ��ϡ�³���褦�Ǥ���
���ܤؤ��ij��������ť롼�Ȥˤ��Ƥ⡢�Ϥ�Ļ�ˤ�봶���Ȥ��Ƥ⡢���줬����뤫���ľ�ԤʤΤ����ڹ��ͳ�γƱ���֤Τ�ΤʤΤ�������Ȥ⡢�ij���ľ���Τ�ΤʤΤ����ޤ����狼��ʤ����Ȥ�¿�����Ǥ��͡�
����顢��衦�����ʩ��������äΤ褦�ˤʤä���ޤ�������
�ij����οʲ��ˤĤ��Ƥϡ���H5N1 Evolution Via Recombination in China�פ���
�ij��г��Ȥ���륦���륹����(2006ǯ6�����)�ϡ�������Υ����ȡ�Qinghai H5N1 Bird Flu Sequences in Bali Indonesia�٤���
�ʤ����ij����ǤϤʤ����ˤ�봶����Ȥ��Ƥϡ��嵭��ʡ�����ˤ�����Ǥδ����Τۤ������Τ�Τ�����ޤ���
�٥ȥʥ�(2004/01)��
����(2004/01)��
����ܥ���(2004/01)��
����ɥͥ���(2004/01)��
�饪��(2004/01)��
�ޥ졼����(2004/05)��
�ߥ��ޡ�(2005/03)
�ޤ�������ʳ���H5N1ȯ����Ȥ��Ƥϡ���ɽ��Ȥ��Ƥϡ����Τ�Τ�����ޤ���
�����åȥ���
1959ǯ��
A/chicken/Scotland/1959
����ꥫ
1966ǯ��
A/turkey/Ontario/7732/66 ��
1975ǯ��
A/Mallard/Wisconsin/428/75 ��
1981ǯ��
A/Duck/Minnesota/1525/81 ��
1983ǯ��
A/Gull/Pennsylvania/4175/83 ����
�����ꥹ
1991ǯ��
A/Turkey/England/50-92/91 ��
���
1996ǯ��
�����A/Goose/Guangdong/1/96 ��
1997ǯ��
����A/chicken/Hubei/wh/1997 ��
1999ǯ��
����������衡A/duck/Guangxi/07/1999 ��
2000ǯ��
���졡A/Goose/Huadong/1/2000 ����ʡ����A/duck/Fujian/19/2000 ��������ʡ�A/duck/Guangdong/07/2000 ������A/duck/Zhejiang/11/2000�ʤɡ�
2001ǯ��
��ǫ��A/Duck/Anyang/AVL-1/2001 �����̡�A/chicken/Hebei/718/2001 �����졡A/chicken/Guangdong/1/2001 ��������A/chicken/Jiande/1218/2001 ��ʡ����A/duck/Fujian/17/2001 ��������²�����衡A/duck/Guangxi/50/2001 ����롡A/duck/Mongolia/54/01-duck/Mongolia/47/01 ���峤��A/duck/Shanghai/08/2001 ��������A/duck/kaifeng/1/01 �ʤɡ�
2002ǯ��
���A/chicken/China/1/02 �����̡�A/chicken/Hebei/108/02 �����ɡ�A/chicken/Jiangsu/cz1/2002�����Ӿʡ�A/chicken/Jilin/hg/2002 ��Ţ����A/chicken/zhengzhou/1/02 ��ʡ����A/duck/Fujian/01/2002�����졡A/duck/Guangdong/22/2002 ��������²�����衡A/duck/Guangxi/53/2002 ���峤��A/duck/Shanghai/35/2002�����ʡ�A/duck/Zhejiang/bj/2002 �����A/quail/yunnan/092/2002 ��A/ostrich/HeNan/14/2002 ������ʤɡ�
2003ǯ
A/Ck/ST/4231/2003 ��A/Dk/HN/5806/2003 �����̡�A/chicken/Hubei/wn/2003 ��A/chicken/Jilin/ha/2003 ��A/chicken/jiyuan/1/03 ��A/chicken/luohuo/3/03 ��A/duck/China/E319-2/03��A/duck/Shantou/4610/2003 ��A/goose/Fujian/bb/2003 ��A/swine/Shandong/2/03 �ʤɡ�
2004ǯ
A/Chicken/Guang xi/12/2004 ��A/Chicken/Hu bei/14/2004 ��A/Ck/YN/115/2004 ��A/Dk/HN/101/2004 ��A/Duck/Guang xi/13/2004 ��A/Ph/ST/44/2004 ��A/babbler/Fujian/320/04 ��A/chicken/China/1204/04 ��A/chicken/Guangdong/174/04 ��A/chicken/Henan/01/2004 ��A/chicken/Macheng/2004 ��A/chicken/Shandong/K01/2004 ��A/chicken/Yichang/lung-1/04 ��A/goose/Huadong/220/2004 ��A/swine/Anhui/ca/2004 ��A/tree sparrow/Henan/1/2004 �ʤɡ�
���
1997ǯ��
A/Chicken/Hong Kong/220/97 �ʤ�
1998ǯ��
A/Hong Kong/97/98 �ʤɡ�
1999ǯ��
A/Environment/Hong Kong/437-10/99 �ʤɡ�
2000ǯ��
A/Duck/Hong Kong/2986.1/2000 �ʤɡ�
2001ǯ��
A/Chicken/Hong Kong/317.5/2001 �ʤɡ�
2002ǯ��
A/Ck/Hong Kong/31.2/2002 �ʤɡ�
2003ǯ��
A/CK/Hong Kong/YU357/03 �ʤ�
2004ǯ��
A/grey heron/Hong Kong/837/2004 ��A/peregrine falcon/Hong Kong/D0028/2004�ʤɡ�
����
2004ǯ��
A/Chicken/Nakorn-Patom/Thailand/CU-K2/2004��A/Chicken/Thailand/73/2004 ��A/Kalji Pheasant/Bangkok/Thailand/CU-18/04 ��A/Prachinburi/6231/2004 ��A/chicken/Kamphaengphet-2-02/2004��A/chicken/Nakhon Pathom/Thailand/CU-14/04��A/chicken/Nakornsawan-2-02/2004 ��A/chicken/Phetchabun-2-01/2004 ��A/chicken/Phitsanulok-2-01/2004 ��A/chicken/Ratchaburi/Thailand/CU-68/04 ��A/chicken/Saraburi/Thailand/CU-17/04��A/chicken/Sukhothai-2-01/2004 ��A/chicken/Uthaithani-2-01/2004 �ʤɡ�
�����ȥʥ�
2001ǯ��
A/Goose/Viet Nam/113/2001 �ʤɡ�
2002ǯ��
A/Duck/Viet Nam/Ncvd1/2002 �ʤ�
2003ǯ��
A/Chicken/Viet Nam/Ncvd8/2003 �ʤ�
2004ǯ��
A/Chicken/Viet Nam/1/2004 ��A/Hanoi/03/2004 ��A/chicken/Viet Nam/AG-010/2004 �ʤɡ�
2005ǯ��
A/Chicken/Viet Nam/NCVD10/2005 �ʤɡ�
����ɥͥ���
2003ǯ
A/Chicken/Indonesia/11/2003 ��A/chicken/Pekalongan/BPPV4/2003 ��A/chicken/Sragen/BPPV4/2003 ��A/chicken/Wonosobo/BPPV4/2003 �ʤɡ�
2004ǯ
A/Ck/Indonesia/4/2004 ��
A/chicken/Bangli Bali/BPPV6-2/2004 ��A/chicken/Denpasar/01/2004 ��A/chicken/Jembrana/BPPV6/2004 ��A/chicken/Kulon Progo/BBVet-XII-1/2004 ��A/chicken/Kupang-1-NTT/BPPV6/2004 ��A/chicken/Malang/BBVet-IV/2004 ��A/chicken/Mangarai-NTT/BPPV6/2004 ��A/chicken/Ngawi/BPPV4/2004 ��A/chicken/Pangkalpinang/BPPV3/2004 ��A/chicken/Purwakarta/BBVet-IV/2004 ��A/chicken/Yogjakarta/BBVet-IX/2004 �ʤɡ�
2005ǯ
A/Chicken/Agam/BBPVI/2005 ��A/Chicken/Deli Derdang/BBPVI/2005 ��A/Chicken/Langkat/BBPV1-576/2005����A/Chicken/Medan/BBPV1-571/2005 ��A/Chicken/Murao Jambi/BBPV-II/2005 �ʤɡ�
����ܥ���
2004ǯ��
A/chicken/Cambodia/1/2004 �ʤɡ�
2005ǯ��
A/Cambodia/JP52a/2005 �ʤɡ�
�ޥ졼����
2004ǯ��
A/chicken/Malaysia/01/2004 �ʤɡ�
�饪��
2004ǯ��
A/Chicken/Laos/44/2004 �ʤɡ�
�٥륮��
2004ǯ��
A/crested eagle/Belgium/01/2004 �ʤɡ�
��եꥫ
2004ǯ��
A/duck/South Africa/811/2004 �ʤɡ�
�ڹ�
2003ǯ
A/Chicken/Korea/es/2003 �ʤ�
����
2003ǯ��
A/duck/Yokohama/aq10/2003
2004ǯ��
A/Chicken/Yamaguchi/7/2004 ��A/chicken/Kyoto/3/2004 ��A/blow fly/Kyoto/93/2004 ��A/crow/Kyoto/53/2004 ��A/chicken/Oita/8/2004 ��A/crow/Osaka/102/2004 ��A/duck/Hokkaido/Vac-1/04�ʤɡ�
���Τ������ǽ��A/duck/Yokohama/aq10/2003;���ͳ����ϡ�2003ǯ5���������ܤ�͢�����줿���ҥ������鸡�Ф��줿��ΤǤ���
��͢�����ҥ�������ο��������ҷ�H5N1Ļ����ե륨�����륹��ʬΥ����
�Ǹ��A/duck/Hokkaido/Vac-1/04�ϡ��褯�狼��ޤ����̳�ƻ��ؤδ��Ķ����δط��Τ褦�Ǥ��͡�
����¾�Υ����륹�Υ������ˤĤ��Ƥϡ���The Influenza Sequence Database�פ���
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/23 Tuesday
2007/01/23(Tue)
��
 �ڹ�������ܤ�Ļ����ե륨�����Ϥ�ŷ�»Ԥ���2����Υ�줿�����ܤ���������ƻ��������������(�ڹ��ǡ�미호천��)�ǡ��ޥ���(�ڹ��Ǥϡ�청둥오리 ��)(Anas platyrhynchos)��ʵ�ʤɤ�ʬ��ʪ����H5N1�����Ф��줿���Ȥǡ���ǯ���Ļ����ե륨�����⡢�Ϥ�Ļ(�ڹ��ǡ�철새��)�ˤ���ΤǤϤʤ��ä��Τ����Ȥ�����¬�����ä�����ޤäƤ��롣
�ڹ�������ܤ�Ļ����ե륨�����Ϥ�ŷ�»Ԥ���2����Υ�줿�����ܤ���������ƻ��������������(�ڹ��ǡ�미호천��)�ǡ��ޥ���(�ڹ��Ǥϡ�청둥오리 ��)(Anas platyrhynchos)��ʵ�ʤɤ�ʬ��ʪ����H5N1�����Ф��줿���Ȥǡ���ǯ���Ļ����ե륨�����⡢�Ϥ�Ļ(�ڹ��ǡ�철새��)�ˤ���ΤǤϤʤ��ä��Τ����Ȥ�����¬�����ä�����ޤäƤ��롣
������ͳ�Ȥ��ơ���ǯ���麣ǯ�ˤ����Ƥ�Ļ����ե륨�����Ϥζ�ˤϡ�������⡢�Ϥ�Ļ���礭�������Ϥ�����Ȥ������Ȥ���
���ʤ������ǯ����켡��ȯ������������������Τ(�ڹ��ǡ�익산시 함열읍 석매리��)�ȡ�����ȯ���������Բ�����(�ڹ��ǡ�익산시��황등면��)�ϡ�������⡢�Ϥ�Ļ�ν�����ϤǤ���ӹ�(�ڹ�����금강��)�Ȥϡ�10������ȥ�;����Υ��Ƥ��餺���軰����ȯ����������Թ�����(�ڹ��ǡ�김제시 공덕면��)�⡢�Ϥ�Ļ�ν�����ϤǤ���������(�ޥ�ڹ�����만경강��)���ܤ��Ƥ��ꡢ��ͼ�ȯ���ϤǤ���绳�������(�ڹ��ǡ�아산시 탕정면��)�⡢�Ϥ�Ļ��¿������ʶ���(�ڹ�����곡교천��)�˶ᤤ��
����������ȯ����ŷ�»ԡ�˭����(�ڹ��ǡ�천안시��풍세면 ��)�����Ȥϡ�˭����(�ڹ��ǡ�풍세천��)��100��ȥ�ε�Υ�ˤ��롣
���Τ褦�ʤ��Ȥ��顢�ڹ����ɤȤ��Ƥϡ��Ϥ�Ļ�δѻ������Ǥʤ�������鴶�������Ϥ�Ļ������ʪ���ܿ������ǽ���Τ�������ȷ�ʤ�αĻ���Ф���ƻ�⡢����빽���Τ褦����
���ȡ�철새 도래지 주변 집중발생 �� 방역당국 긴장��
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/21 Sunday
2007/01/21(Sun) 08:29
��
 �ڹ��������ƻŷ�»ԡʴڹ��ǡ�천안시��)˭���̤��ܷܾ��1��20�������¸���Ļ����ե륨��ȯ������ǧ���줿��
�ڹ��������ƻŷ�»ԡʴڹ��ǡ�천안시��)˭���̤��ܷܾ��1��20�������¸���Ļ����ե륨��ȯ������ǧ���줿��
��ǯ12���������ƻ�绳�ԡ�(�ڹ��ǡ�아산시��)�ǹ��¸���Ļ����ե륨����ǧ����ư��衢5���ܤ�ȯ������
���Υ����ȡ�H5N1 in Central South Korea�פˤ��ȡ����κ�ǯ���麣ǯ�ˤ����ƤΡ��ڹ�����ܤǤ�Ļ����ե륨��ȯ���ξ�������2003ǯ12��� 2004ǯ 1��ˤ����Ƥ�Ļ����ե륨�ξ����ȹ�����Ƥ���Ȥ��Ƥ��롣
���ΤȤ��⡢2003ǯ12��˴ڹ��Ļ����ե륨��ȯ������2004ǯ1������ܤλ����ǡ�Ļ����ե륨��ȯ��������
�����Ǥϡ��ڹ�Ǥ�Ļ����ե륨��ȯ���ϡ�ȯ���������鸫�ơ���뤫�����褷�������ϥ����祦(Whooper swans) (Cygnus cygnus)(�ڹ��ǡ�큰고니��)���ط����Ƥ���ΤǤϤʤ��Ȥ��������Ω�ƤƤ��롣
���͡���������ʡ����Υޥʥť��˿��ϤǤ�Ļ����ե륨������
�����������ܤؤδ����������ϥ����祦�ˤ�ä���ΤȤϡ����ꤷ�ƤϤ��ʤ���
�����ơ����λ����������ϥ����祦�ϡ��ۤȤ�ɡ��������濴�ˤߤ���(�̳�ƻ�����̡����㡢�ٻ�����롦���ϡ����դ��ۤȤ��)�����ޤ�ˡ��彣�Ǥ⡢�����Ѥʤɤ˸���줿�Ȥ����⤢��褦����
���������ڹ�Ǥ�Ļ����ե륨��ȯ���ϤǤ���������ƻ�ʴڹ��ǡ�충청남도�סˤζ���Ϥ�Ļ����©�ϤǤ���������(�ڹ��ǡ�천수만 ��)�ʤɤΥ����ˡ��ǡ�¾��Ļ�˴����������ꡢ�ޥ���ʤɤ��̤��Ϥ�Ļ�������륹�����ܤر�����Ȥβ���⤢�ꤦ���������
 �����ϡ������ѤǤΥ����ϥ����祦�����ͤǤ��롣
�����ϡ������ѤǤΥ����ϥ����祦�����ͤǤ��롣
�ڹ����ƻ�Ǥϡ������������ɱֲ�Ĺ��ȯɽ�ˤ��ȡ�����Ļ����ե륨��ȯ������ŷ�»Ԥ� ����Τ���������용정리산란계농장�˶�2����������˭歲��(풍세천)��������(미호천)��ʬή���Ǻμ褷����Ļ(�ޥ���)(�ڹ��Ǥϡ�청둥오리 ��)(Anas platyrhynchos)��ʬ��ʪ����H5N1�Ф������Ȥ��顢���������ϡ��Ϥ�Ļ(철새)�Ǥ���Ȥθ��������
 �����ϡ�Ʊ�����������Ѥˤ�����ޥ���Υ����ˡ������ͤǤ��롣
�����ϡ�Ʊ�����������Ѥˤ�����ޥ���Υ����ˡ������ͤǤ��롣
������ƻ�϶۵��ɱ��к���Ĥ����Ϥ�Ļ�ܶ�������Ƥ����Ĥ����Ȥ�����
�����Ǥϡ��ָ�����դ��Ϥ�Ļ����θ��������פʤɤΥ���Ƥ��Ф��줿�Ȥ�����
���Υ����������Υ������ϡ������ͻҤ�������ӥǥ��Ǥ��롣(�̿��β��Ρ�300K�ס�56K�פޤ��ϡ��֤��ޡ�����å����Ƥ���������)
�����Ļ����ե륨���Ϥ�Ļ�����⤬���ޤ�ˤĤ졢�����ȯ���Ϥ�ŷ�»Ԥ����ܤ����ǯ����ȯ���ϡ��绳�ԤǤϡ�ͭ�����Ϥ�Ļ�����ϤǤ����곡교천��(���μ̿�����곡교천��)������������ˡ��ҤȤ�����м��ˤʤäƤ���ȤΤ��ȤǤ��롣
���͡�충남도 AI 비상대책회의… 방역 총력 당부 ��
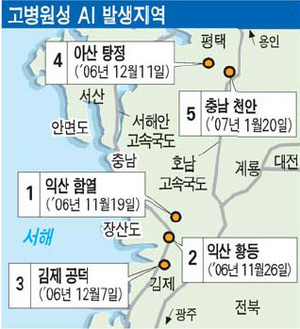

�嵭�οޤϡ���ǯ��11���12��ˤ����Ƥ�Ļ����ե륨��ȯ���ս�ȡ�����ȯ����Ļ����ե륨��ȯ���ΰ��ִط�������Τ���
��천안에서 고병원성 AI 또 발생] 철새가 의심스러워��
��AI, 철새가 옮겼을 가능성 커졌다�פ�����
1.2006ǯ11��19��ȯ����������ƻ�����ԡ�����(�ڹ��Ǥϡ�익산시 함열��)
2.2006ǯ11��26��ȯ����������ƻ�����ԡ�����(�ڹ��Ǥϡ�익산시 황등��)
3.2006ǯ12��07��ȯ����������ƻ������ԡ�����(�ڹ��Ǥϡ�김제시 공덕��)
4.2006ǯ12��21��ȯ����������ƻ���绳�ԡ�����̡��Ȼ�Τ(�ڹ��Ǥϡ�아산시 탕정��)
5.2007ǯ01��20��ȯ����������ƻ��ŷ�»ԡ�˭���̡�ζ��Τ(�ڹ��Ǥϡ�천안시풍년면��)
���ǤΥ����ϥ����祦��H5N1�����ϡ���ǯ��A/whooper swan/Mongolia/2/2006���Υ����륹�Ȥ��ơ���ǧ����Ƥ��롣
���ȡ�H5N1 Qinghai Emergence in Mongolia ��
���Υ��Υ����륹���Ѱۤϡ�����10�ս�ˤ���Ȥ���Ƥ��롣
G59A, T211C, C268T, A539G, C796T, T868C, A1300G, C1330T, G1562T, C1618T.
���Τ������ij���(A/Bar headed goose/Qinghai/1A/2005)�Ǥ��ѰۤȰ��פ��Ƥ���Τϡ��ҤȤĤ�����ơ������Τ褦����
��̣����Τϡ��������ܤεܺ��ȯ�����줿�����륹���Ѱ۲ս꤬�����Υ��Υ����ϥ����祦�Υ����륹A/whooper swan/Mongolia/2/2006���Ѱ۲ս�ȡ��ɤ����ٰ��פ��Ƥ��뤫�ˤ�äơ�������ϩ�����ʤ�Ϥä��ꤷ�Ƥ���ΤǤϤʤ���������
���ʤߤˡ����ܤ����褹�륪���ϥ����祦������Ĵ���ˤĤ��Ƥϡ���������ر�������̿�ʳظ���ʡ����� ��˧������Ρ��������פˤ�ȤŤ����θ��椬����褦����
��Kanai, Y., Sato, F., Ueta, M., Minton, J., Higuchi, H., Soma, M., Mita, N. and Matsui, S. 1997. The migration routes and important restsites of Whooper Swans satellite-tracked from northern Japan��
����1.���Υ����ϥ����祦��H5N1�����륹���Ѱ����ȡ�¾���ϰ�Υ����륹���Ѱ����Ȥ��о�ɽ
��H5N1 Qinghai Emergence in Mongolia �פ�����(�����������Υ����ϥ����祦�Υ����륹)
���͡�Evolution of H5N1 Headlines ��
G59A
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1 DQ343150 A/chicken/Hebei/326/2005 2005 H5N1
DQ320915 A/goose/Shantou/2216/2005 2005 H5N1
AY609312 A/chicken/Guangdong/174/04 2004 H5N1
AY737289 A/chicken/Guangdong/191/04 2004 H5N1
DQ343151 A/chicken/Hebei/718/2001 2001 H5N1
AY585365 A/duck/Guangxi/35/2001 2001 H5N1
T211C
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
DQ676834 A/chicken/Krasnodar/01/2006 2006 H5N1
DQ440535 A/swan/Iran/754/2006 2006 H5N1
DQ412997 A/swan/Italy/179/06 2006 H5N1
ISDN136919 A/swan/Italy/179/2006 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1
C268T
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1 ISDN124038 A/Chicken/Viet Nam/NCVD09/2005 2005 H5N1
ISDN124037 A/Chicken/Viet Nam/NCVD10/2005 2005 H5N1
AM183674 A/chicken/Vietnam/P22/05 2005 H5N1
AM183673 A/chicken/Vietnam/P78/05 2005 H5
DQ497674 A/duck/Vietnam/272/2005 2005 H5N1
AM183676 A/duck/Vietnam/AG40-O2/05 2005 H5
DQ497677 A/mallard/Vietnam/352/2005 2005 H5N1
ISDN111351 A/Chicken/Indonesia/7/2003 2003 H5N1
DQ497646 A/chicken/Sragen/BPPV4/2003 2003 H5N1
DQ497681 A/chicken/Vietnam/28/2003 2003 H5N1
DQ497693 A/chicken/Vietnam/8/2003 2003 H5N1
AY585364 A/duck/Guangxi/22/2001 2001 H5N1
ISDN38260 A/Goose/Viet Nam/113/2001 2001 H5N1
ISDN38261 A/Goose/Viet Nam/324/2001 2001 H5N1
A539G
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
DQ440535 A/swan/Iran/754/2006 2006 H5N1
DQ412997 A/swan/Italy/179/06 2006 H5N1
ISDN136919 A/swan/Italy/179/2006 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1 DQ320939 A/duck/Vietnam/568/2005 2005 H5N1
AY737289 A/chicken/Guangdong/191/04 2004 H5N1
AY518362 A/duck/China/E319-2/03 2003 H5N1
T796C
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1
DQ320939 A/duck/Vietnam/568/2005 2005 H5N1
AY585373 A/duck/Guangdong/07/2000 2000 H5N1
AY059476 A/Duck/Hong Kong/ww381/2000 2000 H5N1
AY059477 A/Duck/Hong Kong/ww382/2000 2000 H5N1
AY059478 A/Duck/Hong Kong/ww461/2000 2000 H5N1
AY059479 A/Duck/Hong Kong/ww487/2000 2000 H5N1
AY059480 A/Goose/Hong Kong/ww491/2000 2000 H5N1
DQ092869 A/Pekin duck/Singapore/F59/04/98 1998 H5N2
AF194992 A/Chicken/Italy/9097/97 1997 H5N9
ISDN49024 A/duck/Singapore/3/97 1997 H5N3
AF303057 A/duck/Singapore/Q-F119-3/97 1997 H5N3
S68489 A/turkey/England/50-92/91 1991 H5N1
AF082042 A/Duck/Potsdam/1402-6/86 1986 H5N2
CY006036 A/duck/Potsdam/2216-4/1984 1984 H5N6
AF082041 A/Duck/Potsdam/2216-4/84 1984 H5N6
M18450 A/Duck/Ireland/113/83 1983 H5N8
M18451 A/Turkey/Ireland/1378/83 1983 H5N8
AF082039 A/Duck/Hong Kong/698/79 1979 H5N3
AF290443 A/Duck/Ho Chi Minh/14/78 1978 H5N3
CY005575 A/duck/Hong Kong/342/1978 1978 H5N2
U20475 A/duck/Hong Kong/342/78 1978 H5N2
CY006028 A/goose/Hong Kong/23/1978 1978 H5N3
AF082038 A/Duck/Hong Kong/205/77 1977 H5N3
AB241615 A/duck/Miyagi/54/76 1976 H5N3
T868C
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
DQ440535 A/swan/Iran/754/2006 2006 H5N1
DQ412997 A/swan/Italy/179/06 2006 H5N1
ISDN136919 A/swan/Italy/179/2006 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1 DQ104701 A/duck broiler/Malaysia/F118/08/04 2004 H5N2
DQ122147 A/duck broiler/Malaysia/F189/07/04 2004 H5N2
AB241624 A/duck/Hokkaido/101/04 2004 H5N3
AB241625 A/duck/Hokkaido/193/04 2004 H5N3
AB241626 A/duck/Hokkaido/299/04 2004 H5N3
AJ715872 A/Hanoi/03/2004 2004 H5N1
AY651335 A/Viet Nam/3046/2004 2004 H5N1
ISDN40278 A/Viet Nam/3212/2004 2004 H5N1
AJ632269 A/chicken/France/03426a/03 2003 H5N2
DQ497679 A/chicken/Vietnam/20/2003 2003 H5N1
DQ320938 A/chicken/Vietnam/27/2003 2003 H5N1
DQ497681 A/chicken/Vietnam/28/2003 2003 H5N1
DQ497693 A/chicken/Vietnam/8/2003 2003 H5N1
AY995885 A/mallard/Sweden/21/02 2002 H5N2
AY995895 A/mallard/Sweden/58/02 2002 H5N3
AF194169 A/Chicken/Italy/312/97 1997 H5N2
A1300G
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1
(plus many other H5N1 isolates)
C1330T
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1
DQ497651 A/chicken/Gunung Kidal/BBVW/2005 2005 H5N1
DQ497643 A/chicken/Magetan/BBVW/2005 2005 H5N1
DQ320933 A/chicken/Wajo/BBVM/2005 2005 H5N1
DQ497659 A/duck/Parepare/BBVM/2005 2005 H5N1
DQ387854 A/mallard/Bavaria/1/2005 2005 H5N2
DQ647648 A/ostrich/South Africa/1/2004 2004 H5N2
DQ007623 A/Anas platyrhynchos/Chany Lake 2003 H5N3
AY995886 A/mallard/Sweden/22/02 2002 H5N9
AY995887 A/mallard/Sweden/27/02 2002 H5N9
AY995888 A/mallard/Sweden/28/02 2002 H5N9
AY995889 A/mallard/Sweden/31/02 2002 H5N2
AY995890 A/mallard/Sweden/37/02 2002 H5N9
AY995891 A/mallard/Sweden/39/02 2002 H5N3
AY995892 A/mallard/Sweden/40/02 2002 H5N6
AY995893 A/mallard/Sweden/49/02 2002 H5N9
AY995894 A/mallard/Sweden/52/02 2002 H5N9
AY995897 A/mallard/Sweden/79/02 2002 H5N9
AY995898 A/mallard/Sweden/80/02 2002 H5N9
AJ621807 A/duck/Primorie/2633/01 2001 H5N3
AY075030 A/Goose/Hong Kong/3014.5/2000 2000 H5N1
AY059482 A/Goose/Hong Kong/3014.8/2000 2000 H5N1
AY684894 A/mallard/Netherlands/3/99 1999 H5N2
AJ305306 A/chicken/Italy/8/98 1998 H5N2
AF194169 A/Chicken/Italy/312/97 1997 H5N2
AF194990 A/Chicken/Italy/367/97 1997 H5N2
AF194992 A/Chicken/Italy/9097/97 1997 H5N9
AF194991 A/Guinea Fowl/Italy/330/97 1997 H5N2
G1562T
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1 DQ497642 A/chicken/Malang/BBVet-IV/2004 2004 H5N1
DQ497653 A/chicken/Purwakarta/BBVet-IV/2004 2004 H5N1
C1618T
DQ661910 A/chicken/Afghanistan/1207/2006 2006 H5N1
ISDN133364 A/Turkey/65596/06 2006 H5N1
AB263752 A/whooper swan/Mongolia/2/06 2006 H5N1
DQ320910 A/chicken/Hunan/999/2005 2005 H5N1
DQ095630 A/Duck/Hunan/114/05 2005 H5N1
DQ320902 A/duck/Hunan/127/2005 2005 H5N1
DQ320903 A/duck/Hunan/139/2005 2005 H5N1
DQ320904 A/duck/Hunan/149/2005 2005 H5N1
DQ320905 A/duck/Hunan/152/2005 2005 H5N1
DQ320906 A/duck/Hunan/157/2005 2005 H5N1
DQ320907 A/duck/Hunan/160/2005 2005 H5N1
DQ320908 A/duck/Hunan/166/2005 2005 H5N1
DQ320909 A/duck/Hunan/182/2005 2005 H5N1
DQ095631 A/Duck/Hunan/191/05 2005 H5N1
DQ387854 A/mallard/Bavaria/1/2005 2005 H5N2
AF509025 A/Chicken/Hong Kong/715.5/01 2001 H5N1
AY221524 A/Chicken/Hong Kong/FY150/01 2001 H5N1
AF509019 A/Chicken/Hong Kong/FY150/01 2001 H5N1
AY221523 A/Chicken/Hong Kong/FY150/01-MB 2001 H5N1
AF509016 A/Chicken/Hong Kong/FY77/01 HA 2001 H5N1
AY221522 A/Chicken/Hong Kong/NT873.3/01 2001 H5N1
AY221521 A/Chicken/Hong Kong/NT873.3/01-MB 2001 H5N1
AY221526 A/Pheasant/Hong Kong/FY155/01 2001 H5N1
AF509020 A/Pheasant/Hong Kong/FY155/01 2001 H5N1
AY221525 A/Pheasant/Hong Kong/FY155/01-MB 2001 H5N1
M18450 A/Duck/Ireland/113/83 1983 H5N8
M18451 A/Turkey/Ireland/1378/83 1983 H5N8
X07869 A/chicken/Scotland/59 1959 H5N1
X07826 A/Chicken/Scotland/59 1959 H5N1
����2.���ij������Ѱ۲ս�
�������̤ꡣ
RNA�ե饰����(RNA fragments)1.PB2
A1398G T1460C��
RNA�ե饰����2.PB1
T87C C557T G1362T T2196C��
RNA�ե饰����3.PA
T147C G228A T571C T1594C G1825G��
RNA�ե饰����4.HA
G59A�� C796T A1300G��G1562T��
RNA�ե饰����5.NP
C74T G343A ��
RNA�ե饰����6.NA
C162T C979T G1361A��
RNA�ե饰����7.MP
C559A��
RNA�ե饰����8.NS
T41A A849G A851G��
G59A, T211C, C268T, A539G, C796T, T868C, A1300G, C1330T, G1562T, C1618T.
���ȡ�Tuva and Mongolia H5N1 Bird Flu Sequences Are Similar��
A/whooper swan/Mongolia/2/2006�ΰ���������ˤĤ��Ƥϡ����Υ�����������
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/19 Friday
2007/01/19(Fri)
��
 �ȣ��Σ��˴�������ǯ���˻�˴���������ץȤΣ��ͤ��顢�������륹�����ߥե�������Τ��륦���륹�����Ф��줿�ȡ������ݷءʣףȣϡˤ�������ȯɽ�����Ȥ����Τ�����
�ȣ��Σ��˴�������ǯ���˻�˴���������ץȤΣ��ͤ��顢�������륹�����ߥե�������Τ��륦���륹�����Ф��줿�ȡ������ݷءʣףȣϡˤ�������ȯɽ�����Ȥ����Τ�����
���ȡ�Avian influenza - situation in Egypt ��
�ףȣϤˤ��ȡ������ץ�����Gharbiya��Ʊ���Ȥ˽���Ƥ���Ʊ�Τ�Warda Eidh Ahmed�Ȥ���̾�Σ����н����ȣ������������꼡���Ǵ������������Τۤ��ϡ���ǯ12��19���ˡ����������ϡ���ǯ12��17������������12��21�����顢����2���Υ��ߥե�ˤ�뼣�Ť�Ϥ�Ȥ�����
12��23���ˡ���ͤϡ�referral hospital�ذܤ��줿���������Τۤ��ϡ�12��25���ˡ����������ϡ�12��28���ˡ���˴�����Ȥ�����
��Ϳ���ϣ�����˺μ褷�����ΤΡ��ݥ������294��N����S �ؤ��Ѱ�(N294S)������졢���ߥե�������٤������Τ��륦���륹�Ǥ��뤳�Ȥ��狼�ä��Ȥ�����
������������Ȥ��äˤ��ȡ����ߥե�μ��Ť�Ϥ�Ƥ����Ѱۤˤ�����ޤǤλ��֤�û������ΤǤϤʤ����Ȥλ�Ŧ�⤢�롣
�����ǡ��ºݤϡ��ij����ˤ����Ƥ⡢Ʊ��N294S�Ѱۤ����ä��ˤ⤫����餺�����ˤ���ʤ��ä��ΤǤϤʤ����ȡ���¬��������⤢��褦����
�Ĥޤꡢ�����ץȤǥ��ߥե뼣�Ť�Ԥ������顢�����륹�ˤϡ����ߥե�����������äƤ����ΤǤϤʤ����Ȥ�����¬����
N294S�ϡ�Vietnam�Ǥ�ߤ�줿���ߥե������ѰۤΤҤȤĤǤ⤢�ꡢ�����ץȤǤΥ��ߥե����Ѥˤ���ѰۤǤϤʤ��褦�ˤ�ߤ��롣
�ޤ����ҥȤؤδ����ȴط��Τ���receptor binding domain�Υݥ������230�Ǥ�M����I�ؤ��Ѱ�(M230I)��ǧ���줿�Ȥ�����
M230I���Ѱۤϡ��ҥȡ�����ե륨�� H1N1, H3N2, ����ե륨�� B�λ��ĤΥ����פˤ⸫�����ѰۤǤ��ꡢ�����.Ļ����ե륨�����륹���ѰۤǤϤʤ���HIV���Ѱۤ��Ȥ��븫���⤢��褦����
�ݥ������230��M����I�ؤ��Ѱ�(M230I)������줿�������Ȥ��Ƥϡ����ǡ�2001ǯ��2002ǯ�˸���줿�����륹�����(A/duck/Zhejiang/bj/2002(H5N1) �ȡ�A/Duck/Hong Kong/380.5/2001)�����롣
���Υ����ץȤδ��ԤΥ����륹�˸������ѰۤȤ��Ƥϡ����Τۤ��ˡ�M230I��Ʊ�����������η��ɥᥤ��Ǥ��ѰۤȤ��ơ�V223I��������Ȥ�����
V223I�ϡ�A/bar-headed goose/Mongolia/1/05�ˤ⡢A/chicken/Yamaguchi/7/2004�ˤ⸫��줿�ѰۤǤ��롣
�ʤ������ߥե������Υ����륹���ѰۤȤ��Ƥϡ�H5N1�Ǥϡ�R152K��H274Y��N294S�����ꡢH5N2�Ǥϡ�E119G/A/D/V,R292K�����롣
H5N1
R152K(�ݥ������152�ǤΥ��륮�˥�ꥷ��ؤ��Ѱۡ�N1)
H274Y(�ݥ������274�ǤΡ��ҥ��������������ؤ��Ѱۡ�N1) ��
N294S(�ݥ������294�ǤΥ����ѥ饮�饻���ؤ��Ѱۡ�N1)
H5N2
E119V��G��A��D(�ݥ������119�ǤΡ����륿�ߥ������Х��ޤ��ϥ��ꥷ��ޤ��ϥ���˥�ޤ��ϥ����ѥ饮����ؤ��Ѱۡ�N2 ),
R292K(�ݥ������292�ǤΥ��륮�˥�ꥷ��ؤ��Ѱۡ���2)
������Τ�����H274Y(�ݥ������274�ǤΡ��ҥ��������������ؤ��Ѱۡ�N1)�ˤĤ��Ƥϡ������ȥʥ�ξ����ǥ��ߥե����������ä���Τǡ�����ˤĤ��Ƥϡ���ΰ����Υ֥���������ե륨��A��H5N1����������Υ��ߥե������ˤĤ����פ˾ܤ��������Ȥ����롣
�ʤ���HIV��H5N1�Ȥδط��ˤĤ��Ƥϡ������ȡ�HIV Could Launch Bird Flu��Pandemic �� �ˤ����ơ����Τ褦�˽�Ƥ��롣
Robert Webster ��Τˤ��С����Ǥ�HIV�˴������Ƥ��봵�Ԥϡ��ȱַϤ���äƤ��뤿��ˡ�H5N1�ʹ֤˴������䤹�������Ѥ��Ƥ��ޤ��Ȥ�����
����ϡ��ʤˤ⡢HIV�˸¤餺��Ʊ�������ȱַϤ���äƤ��롢���ԤˤĤ��Ƥ������ȡ�Robert Webster ��Τϡ����äƤ��롣
���ʤߤˡ������ץȤˤ�����HIV���Ԥϡ������ˤϡ�1986ǯ�衢1800�ͤȤ������Ȥ������ºݤˤϡ�����ʳ��˱����줿���Ԥ�����������ȿ�¬���졢���ο��ϡ�2003ǯ�����ǡ�5000�ͤ���30000�ͤ�������Ȥ��졢����Ǥ����ȡ������ץȿͤ�2355�ͤ˰�ͤ���HIV�Ǥ���Ȥο����⤢��褦����
���͡�Egypt confronting HIV��
�ޤ���Ʊ�������Ȥο����Ǥϡ�����ɥͥ����ˤ�����HIV�ϡ�130000�ͤȤ����Ƥ��롣
��������HIV��¿����Ǥ�H5N1�ˤ���˴�Ԥ�¿���Τ������ˤʤ�Ȥ�������
����
��N294S Tamiflu Resistance Marker In H5N1 Ducks�פʤ�
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/12 Friday
2007/01/12(Fri)
��
 �ܺ긩�ܺ귴����Į�ڸ��Ρ�ë���������������פǺ���������飱�����ˤ����ơ����餷�Ƥ���˥�ȥ꣱�����鱩�Τ��������褽����������������
�ܺ긩�ܺ귴����Į�ڸ��Ρ�ë���������������פǺ���������飱�����ˤ����ơ����餷�Ƥ���˥�ȥ꣱�����鱩�Τ��������褽����������������
���θ�⡢�����ˡ��ܤ����Ǥ��ꡢ���������������������������������������������2,400��(Ʊ�ܷܾ�����������������Σ�ʬ�Σ�)��˴�ȤʤäƤ��롣
�ð�դˤ��ʰ����Ǥϡ�Ļ����ե륨�����륹������ȿ�����ФƤ��ꡢ�ܺ긩�����ݷ�����꤬����˥����륹ʬΥ������»ܤ�����븩�Ĥ��лԤ�ưʪ���������θ�����Ф��塢��̤�13����Ƚ������ȤΤ��ȤǤ��롣(2007/01/13�ɵ����ܺ긩�ˤ�����Ļ����ե륨�����륹�ϡ��ȣ������Σ�������ե륨�Ǥ���Ȥ�ȯɽ�����ä�����N1�ʤΤ��ɤ������䡢�����ҷ��ˤĤ��Ƥ�ȯɽ�ϡ������ۤ��줿�褦����
2007/01/16�ɵ���ưʪ����������ȯɽ�ˤ��ȡ����ܺ긩����Į��Ļ����ե륨�Υ����륹�ϡ��ȣ��Σ����Ǥ��뤳�Ȥ���ǧ���줿�������Ҳ��Ϥ�ȯɽ�Ϥޤ��Τ褦����)
���������ܺ긩�ϡ������ܷܾ���Υ����ȤȤ�ˡ�Ⱦ�£�����������ˤ��룱�����ܷܾ�ˤ�˥�ȥ����ʤɤΰ�ư�ͤ���褦����������
1.����Į���դ��Ϥ�Ļ������
�ϿޤǸ���¤�����������Į�ˤϡ��äˤ������ʿ�Ļ�����Ϥϡ��ʤ��褦�ʤΤ�����������䡢�����ؤβϸ��ˤ϶ᤤ�褦����
������εܺ긩¦�ˤϡ��������ϸ���(�����ܺ��)�䡢�ܺ���ΰ�������깾(�ܺ��)��Ϥ���Ȥ��ơ�¿���δ���ʤɤ����ꡢ�����ˤϡ��߾�Ǥϡ��������������復�����륬�⡢�ޥ��⡢�ߥ�������������������������ᡢ�������ɥꡢ�ϥ��ܥ����饹�����������ʤɤο�Ļ��¿��������褦����
�������ϸ���ϡ��ڹ�κѽ���ʤɤȤȤ�ˡ������ĥ�إ饵���α����ϤȤ��Ƥ⡢�Τ��Ƥ���褦����
�������ο�Ļ�ˤĤ��Ƥϡ����Υ�����
��������깾�ο�Ļ�ˤĤ��Ƥϡ����Υ�����
������
2.�Ϥ�Ļ�ˤ�륦���륹���Ťβ�ǽ��
���λ����δڹ�Ȥ��Ϥ�Ļ�α���ˤĤ��Ƥϡ��狼��ʤ�������ǯ11�����δڹ��Ļ����ե륨��ȯ���Ȥδط��ϡ����Ƥ����ʤ��Ȥ����ʤ���������
 �Ĥޤꡢ�����οޤδڹ�Υե饤���������������ˡ����ܤζ彣������Ȥ������¤���
�Ĥޤꡢ�����οޤδڹ�Υե饤���������������ˡ����ܤζ彣������Ȥ������¤���
����ˤĤ��Ƥϡ�����̤Υ֥����������ڹ�ǡ��絬�Ϥ�Ļ����ե륨��ȯ����������
���ʤߤˡ��ܺ긩�ϡ������ܷ������ʤɡ��֥����顼�������������ϤǤ��ꡢ���ȵܺ귴�Ȥϡ��ܺ�Ԥ�֤˶���ǡ�������褤�ˡ��٤��碌�ˤʤäƤ��롣
�����Υ����륹���ɤΥ롼�ȤǤ������������Ϥ�Ļ�ˤ�������β�ǽ����������
.
.
3.�������̤�.�ּ����η��ɥᥤ��פΰ�����������Ѱۤǡ��Ϥ�Ļ���������������������١�����Ǥ���-�ij�����?ʡ������?-
�Ϥ�Ļ������ˤ�ä���硢���γ�������( cleavage site ��Amino acid at position 323-330)�ΰ��������������Τɤ���˳������뤫�ˤ�äơ����������������ʤ����Τˤʤ��������
���͡�H5N1 cleavage sites ��
���ʤߤˡ��ڹ��Ļ����ե륨�����륹�ϡ��������ܡ֡�H5N1 �ץ����륹�γ������̤�PB2 E627K����������15�Υѥ�����פΤ����έ����ij���(the Qinghai strain)��Ʊ���Ǥ��ä���
���ȡ�Qinghai H5N1 Confirmed On South Korean Farms��
�ij���(the Qinghai strain)�ϡ�2005ǯ5��ˡ������ij��Ф�ȯ�����줿��Τǡ��������顢2005ǯ�Ƥˡ�����������롢�����ե�����ˤ錄�ꡢ���줫�顢2005ǯ��Ⱦ����2006ǯ��Ƭ�ˤ����ơ��衼���åѡ����졢���եꥫ������ɡ����ե��˥�����ؤȡ����礷�Ƥ��ä���ΤǤ��롣
���Υ����ȡ���Suspect Qinghai H5N1 Infections in Japan�פǤϡ����ܤΤ�Τ⡢�����ij�����the Qinghai strain�ˤǤ������ȿ�¬���Ƥ��롣
����Ȥ��̤ˡ�2005ǯ3�������ʡ���ʤǸ���졢���θ塢������饪�����ޥ졼�����������ʤɡ���������˸���줿A/DK/Fujian/1734/05(H5N1)�ʤɡ�ʡ����(the Fujian strain )�ȸƤФ���Τ����뤬�����Τʤ��ǡ�A/Guinea fowl/Shantou/1341/2006(H5N1)���������ij�����the Qinghai strain�ˤǤ���Ȥ���Ƥ��롣
����ˡ�������5�ǽҤ٤�ּ����η��ɥᥤ��פΰ�����������Ѱۤˤ�äƤ⡢�ɤ���ȯ�����줿�����륹�ʤΤ������������١�����Ǥ��롣
����
��ʡ��������Ļ����ե륨���礭���װ��ȤΥ���ꥫ�������Ʊ���������λ�Ŧ��
��Study says new H5N1 strain pervades southern China��
4.Ļ����ե륨�������륹�γ������̤�PB2 E627K����������ǡ�����������ȥ������ȡ����⤿�餹
 ����ޤ�ȯ������Ļ����ե륨�������륹�γ������̤�PB2 E627K����������Ƥߤ�ȡ������Υѥ������ʬ����줦�롣
����ޤ�ȯ������Ļ����ե륨�������륹�γ������̤�PB2 E627K����������Ƥߤ�ȡ������Υѥ������ʬ����줦�롣
���ʤߤˡ�����PB����� E627K����ʬ���Ѱۤ�������ȡ������ȥ������ȡ���(�ȱֱ����ΰ۾�ˤ�륵���ȥ�����β�껺��)ͶƳ��⤿�餹��ǽ�����⤤�Ȥ���롣
���͡�Fujian H5N1 Bird Flu Spreads Throughout China ��
��China releases human H5N1 sequences��
��H5N1 cleavage site options��
��Novel Human H5N1in Indonesia Raises Pandemic Concerns��
��Flu Wiki 2 - Forum - RESRRKKR��
��Growing H5N1 Transmission Chain in Indonesia Raises Concerns ��
��Evolution and Spread of Fujian H5N1 In China��
��A list of relevant mutation position ��
��H5N1 HA Cleavage Site Evolution ��
��Doctors find more evidence bird flu can evade drug��
��Qinghai HA in Indonesia��
��H5N1 cleavage site options��
��Important H5N1 Treatment Hints��
��Example 1: 453 avian influenza HA genes��
(1)��H5N1 �ץ����륹�γ������̤�PB2 E627K����������Υѥ�����
��-���Υ������θ����Ǥ�����������amino acid at position 323����Ϥޤ�amino acid at position 330�ޤǤ�7�ĤΥݥ������ˤʤäƤ��ޤ���
����ե��٥åȤμ�����̣�Ǥ��������ΤȤ���Ǥ���
R=���륮�˥�Arginine (Arg) E=���륿�ߥ��Glutamic Acid (Glu) S=�����Serine (Ser) K=�ꥸ��Lysine (Lys) G=���ꥷ��Glycine (Gly) I=������������Isoleucine (Ile) D=�����ѥ饮���Aspartic Acid (Asp) Q=���륿�ߥ�Glutamine (Gln)
����˾ܤ����ϡ�������Υ����ȡ�The Single-Letter Amino Acid Code�פΰ���ɽ��ɤ�����
����RESRRKKR��-����ɥͥ���(2005ǯ��2006ǯȯ��)������ɥͥ�����ǭ���Τ�Ρ�(A/Indonesia/321H/2006�ʤ�)
����RERRRKKR��-�������Ǥ�äȤ�ݥԥ�顼�ʥѥ�����1997ǯ�����2004ǯ�����ȥʥ��ȯ���������ȥʥࡢ����������ܥ���������(A/ Viet Nam/ 1194/ 2004�ʤ�)
����RERRKKR��-��������ǯ�Υ����ȥʥ�ˤ�����������̤�����ǡ��嵭�������R�פ��ҤȤ�̵���ʤäƤ��룲������ǯ�����ȥʥ������(���ܤ�A/chicken/Yamaguchi/7/04�ʤ�)
����RERRRKR��-���ʡ���������Υ����륹�ǡ��嵭�����顢��K�פ��ҤȤĤʤ��ʤꡢ��R�פ��ҤȤĤդ��Ƥ��롣(A/DK/Fujian/1734/05�ʤ�)
����GERRRKKR��-�����ij��ФǤ�ȯ�������륹�ǡ�������Ƭ�Ρ�R�פ���G�פ��Ѥ�äƤ��롣����������ڹ���������롢�����ե����衼���åѡ����졢���եꥫ������ɡ����ե��˥����ȥ륳�����饯��������Х���������ץȤΥ����륹������⤳���Ʊ��������ɥͥ����ΥХ����ȯ�����줿A/duck/Badung-Bali/05/2005��Ʊ��(Goose/Qinghai/0510/05��A/chicken/Egypt/2253-1/2006�ʤ�)
����REGRRKKR��-�����( A / swine / Anhui / 2004�ʤ�)
����REGGRRKR��-������2(A/chicken/Hunan/2246/2006�ʤ�)
����RERRRRR��-������3����ͤοʹ֤�긡��(A/Hunan-Xiangtan-he/12/2005�ʤ�)
����GERRRRKR��-������Ļ��긡��(A/whooper swan/Mongolia/3/05�ʤ�)
����REGGRKKR��-�����(A/chicken/Hebei/326/2005�ʤ�)
����RERRRIKR & RERRRIKK���ݥ����(2003ǯ)
����RDRRRKKR������峤(���ҥ�)(A/duck/Shanghai/1/2000�ʤ�)
����GEGRRKKR���ݥ�������(A/chicken/Sudan/1784-10/2006�ʤ�)
����GDRRRKKR���ݥ롼�ޥ˥�(A/chicken/Romania/4793/Tl/2005�ʤ�)
����KERRRKKR���ݥ����ץȡ�����(A/chicken/Egypt/5610NAMRU3-F3/2006��A/Thailand/HA20/2005)
����GKRRRKKR���ݥ����ץ�(A/chicken/Egypt/5611NAMRU3-AN/2006)
����GERIRKKR, GERKRKKR, GEKRRKKR��-novel Europe
(��H5N1 cleavage sites ��
��H5N1 cleavage site options - FluTrackers��
��Cleavage��
��The Single-Letter Amino Acid Code����)
(2)ʡ�����ˤ����볫�����̤ξ�ή������ΰ㤤
ʡ�����ˤ����Ƥϡ��������̤ξ�ή�Υ������Ǥ���amino acid at position 315(314)�Υ���������¾�Υ��������ȡ����Τ褦�˰ۤʤäƤ��롣
ʡ�����Ȥ���¾�Υ��������Ȥ�����ΰ㤤
ʡ����������-��LRERRRKR�Ͱʲ����������̤Υ���������Ƭ�Υݥ������(323)�Ǥ����R�פˤĤʤ��äƤ�������
�������˸���������-��QRERRRKKR�Ͱʲ����������̤Υ���������Ƭ�Υݥ������(323)�Ǥ����R�פޤ��ϡ�G�פˤĤʤ��äƤ�������
ʡ�����Ǥϡ�������������Ρ�Q�פ���L�פ��Ѱۤ���K�פ�����Ƥ��롣
����
��Evolution and Spread of Fujian H5N1 In China��
��Dr Niman on cleavage site evolution ��
��Engineered viral vaccine constructs with dual specificity: Avian influenza and Newcastle disease��
��H5N1 Confirmed On South Korean Farm��
5.�ּ����η��ɥᥤ��פǤ��Ѱۤ����ͤؤδ������ưפ�������դ��롣
H5N1��receptor-binding domain (RBD)(���ܸ�Ǥϡ��ּ����η��ɥᥤ��פȤǤ������ΤǤ��礦����)�Ǥ��Ѱۤ����إޥ�����˥��ҥȼ����Τ˷�礷�פ������ͤؤδ������ưפ������š�����Ȥ���Ƥ��롣
����receptor-binding site�ϡ��ȣ��ˤ�äưۤʤꡢ
H3��numbering system��Ĥ��äƤߤ�(H5�ξ��ϡ��ݥ������ʥ�С���������ȰۤʤäƤ���Τǡ������ա�)�ȡ�
amino acids position �Ρ�98, 134–138, 153, 155,183, 190, 194, 195��224–229 �ʤɤȤ���Ƥ��롣
���Τ�����position 190�ޤǤ��ή(upstream)��position 190�ʹߤ�ή(downstream)�Ȥ��Ƥ���褦����
���� receptor binding domain �ˤ���������ҤΥ����������ͤؤδ�����������ΤȤ���Ƥ��롣
���ȡ�The index influenza A virus subtype H5N1��isolated from a human in 1997 differs in its��receptor-binding properties from a virulent��avian influenza virus��
��Qinghai H5N1 Receptor Binding Domain Changes In Middle East��
��H5N1 Acquisition Matches Influenza B Receptor Binding Domain��
����ޤ��Ѱۤ�����줿 receptor binding domain �ϡ������ΤȤ���Ǥ��롣
�ʤ������θ����Ǥ��������Ȥ��С���D187N �פȽƤ�����ˤϡ����ݥ������187�ˤ����ơ�D(�����ѥ饮���)����N(�����ѥ饮��)���Ѱۤ����פȤ������Ȥ�ؤ��ޤ���
���ߥλ�����ˤĤ��Ƥϡ����Υ����ȡ�DNA �ȥ��ߥλ��ΰŹ�ɽ �פ��Ȥ���������
Amino acid residue position 190�ξ�ή(upstream)��
amino acid at position��186(position186�ǤΥ��ߥλ������Ѱۡ��ʲ�Ʊ��)-
N186K(QinghaiʬΥ����AzerbaijanʬΥ��)(�����ѥ饮��ꥷ����Ѱ�)
or
N186S(QinghaiʬΥ��)(�����ѥ饮�饻�����Ѱ�)
amino acid at position 187(position187�ǤΥ��ߥλ������Ѱۡ��ʲ�Ʊ��)-
D187N (HunanʬΥ��-isolates-��A/chicken/Hunan/2246/2006�ʤ�) (�����ѥ饮������饢���ѥ饮����Ѱ�)
amino acid at position 188-
A188E (ShanxiʬΥ����A/chicken/Shanxi/2/2006�ʤ�) (����˥饰�륿�ߥ�����Ѱ�)
amino acid at position 189-
A189T (ShanxiʬΥ����A/chicken/Shanxi/2/2006�ʤ�) (����˥饹�쥪�˥���Ѱ�)
or
A189E (HunanʬΥ����A/chicken/Hunan/2246/2006�ʤ�) (����˥饰�륿�ߥ�����Ѱ�)
Amino acid residue position 190�β�ή(downstream)
amino acid at position 190-
E190D��( VietnamʬΥ����A/Vietnam/1203/2004�ʤ�) (���륿�ߥ�����饢���ѥ饮������Ѱ�)
or
E190V ( VietnamʬΥ��) (���륿�ߥ������Х����Ѱ�)
amino acid at position 192-
T192I (Shanxi ����Hunan��ʬΥ����A/chicken/Shanxi/2/2006��A/chicken/Hunan/2246/2006�ʤɡ�) (���쥪�˥饤������������Ѱ�)
amino acid at position 194-
L194I (Shanxi ����Hunan��ʬΥ����A/chicken/Shanxi/2/2006����A/chicken/Hunan/2246/2006�ʤ�) (�������饤������������Ѱ�)
amino acid at position��196-
Q196R(QinghaiʬΥ��)(���륿�ߥ饢�륮�˥���Ѱ�)
amino acid at position 214-
V214M (Shantou ʬΥ����A/goose/Shantou/2086/2006�ʤ�) (�Х�������˥���Ѱ�)
amino acid at position 220-
R220K (Shanxi ʬΥ������A/chicken/Shanxi/2/2006�ʤ�) (���륮�˥�ꥷ����Ѱ�)
amino acid at position 222-
K222Q (Shanxi ʬΥ����A/chicken/Shanxi/2/2006�ʤ�) (�ꥷ�饰�륿�ߥ���Ѱ�)
or
K222R (Shantou ʬΥ����A/goose/Shantou/2086/2006�ʤ�) (�ꥷ�饢�륮�˥���Ѱ�)
amino acid at position 223-
V223I (Shantou ʬΥ����A/goose/Shantou/2086/2006�ʤ�) (�Х�饤������������Ѱ�)
amino acid at position 225-
G225D ( VietnamʬΥ����A/Vietnam/1203/2004�ʤ�) (���ꥷ�饢���ѥ饮������Ѱ�)
amino acid at position 226-
Q226L ( VietnamʬΥ����A/Vietnam/1203/2004) (���륿�ߥ����������Ѱ�)
amino acid at position 227-
S227R (Shantou ʬΥ������QinghaiʬΥ������A/goose/Shantou/2086/2006�ʤ�) (����饢�륮�˥���Ѱ�)��
or
S227N��Turkey��& VietnamʬΥ���� A/turkey/Kfar Warburg/1224/03 A/Vietnam/1203/2004�ʤɡ����γ��ϡ�H9N2 ��H5N1�Ȥ��ߥå���������ΤȤ����Ƥ��롣�� (����饢���ѥ饮����Ѱ�)
amino acid at position 228-
G228S��( VietnamʬΥ�� A/Vietnam/1203/2004 �ʤ�) (���ꥷ�饻�����Ѱ�)
amino acid at position 230-
M230I ��EgyptʬΥ����QinghaiʬΥ����ZhejiangʬΥ����A/Egypt/12374-NAMRU3/2006��A/duck/Zhejiang/bj/2002(���ʤɡ��� (������˥饤������������Ѱ�)
���͡�H5N1 Fujian Cleavage Site Evolution��
��OIE Hong Kong Report ��
��Hunan H5N1 Has Multiple Receptor Binding Domain Changes ��
��A list of relevant mutation position ��
��H5N1 genetic structure��
��If anyone one is interested in the progress of H5N1 from sources doing histology ��
��Qinghai H5N1 Receptor Binding Domain Changes In Middle East��
�ʤ����ʲ����ֳ��Ǥ��뤬���ҥȴ���H5N1�����륹�ǡ����ߥե����Ѥˤ�äƥ��ߥե����������������ѰۤȤ��Ƥϡ������Τ�Τ����롣
R152K(�ݥ������152�ǤΥ��륮�˥�ꥷ��ؤ��Ѱۡ�N1)( B/Memphis/20/96�����ʤ�)
H274Y(�ݥ������274�ǤΡ��ҥ��������������ؤ��Ѱۡ�N1) (A/Texas/36/91�ʤ�)��
N294S(�ݥ������294�ǤΥ����ѥ饮�饻���ؤ��Ѱۡ�N1) �� Qinghai���ˡ������Ѱۤ�����줿�Ȥ���Ƥ��뤬��������ȯɽ�Ϥʤ��褦������
�ʤ������Υ����ȡ�Seasonal & Pandemic Influenza�פˤ��С����ܤξ�硢E119V,R292K,N294S���Ѱۤϡ�H3N2�����륹���Ф��ơ�18�ѡ�����Ȥξ�������������H274Y���Ѱۤϡ�H1N1�����륹���Ф��ơ�18�ѡ�����Ȥξ��������������Ȥ���롣
�����륹���Ȥι��ʤ�ܺ٤ʰ���������ˤĤ��Ƥϡ���The Influenza Sequence Database�פ���
���Ȥ��С�����ˡ������륹̾��A/Bar headed goose/Qinghai/1A/2005 �פ����Ϥ���ȡ�Raw Sequence�������롣
����ˤ�ä�����줿��Raw Sequence�פ��amino acids������Ǹ���ˤϡ�����ˡ���Sequence Manipulation Suite�פ��Translation of DNA to an Amino Acid Sequence�פ��BCM Search Launcher�פ��Translate a DNA Sequence�פ��Translation of DNA to an Amino Acid Sequence�פʤɤ�Translation�ˤ�����ɬ�פ����롣
��Raw Sequence�פ�Translation�ˤϡ���Geneious �פʤɡ��ե���եȤΥ���������ɤˤ�äƤ⡢��ǽ�Ǥ��롣
6.���ܤ�Ļ����ե륨���說����Υ��졼��2�б��ϡ��Ϥޤä��Ф���
�ʤ������ߤ�H5N1�Υ說�����б��ϡ���ȿ�����ˤ������äơ�����ʬ��ˤ�륯�졼��(����ʬ��)�б��ȤʤäƤ��롣
���졼�ɤ��Ȥΰ���������ΰ㤤�ϡ����Υ�������Evolution of H5N1 avian influenza viruses in Asia�פ���
�����졼��1
-�����ȥʥ���濴�Ȥ����������
�����졼��2
���֡����졼�ɭ�
-����ɥͥ���
���֡����졼�ɭ�
-�ij���������������ڹ���������롢�����ե����衼���åѡ����졢���եꥫ������ɡ����ե��˥�����
���֡����졼�ɭ�
-ʡ�����������������������饪�����ޥ졼�����������ʤɤ��������
���ȡ�Qinghai H5N1 Confirmed On South Korean Farms��
����ޤǡ����ܤ�H5N1�說����ϡ��٥ȥʥ�����Ȥˤ������졼��1�б��Ȥ��Ƥ���������ǯ���ˡ����졼��2�б��說����Ȥ��ơ�����ɥͥ��������Ȥˤ������졼��2���б��說����ȯ���ᡢ��Ω�����������б��ѤߤǤ��롣
����Ū�ˤϡ����졼��2�ϡ��嵭�Τ褦�ˡ�����ˡ����֥��졼�ɭ����������˺�ʬ�������б�����褦�ˤʤäƤ��롣
��ǯ�������ܤΥ��졼��2�б��說����ϡ����Τ����Υ��֥��졼�ɭ��б��說����Τ褦�Ǥ��롣
�����������줬�����졼��2�����֥��졼�ɭ��ȡ����졼��2�����֥��졼�ɭ��ˤĤ��Ƥ�ͭ���Ǥ��뤫�ɤ����ˤĤ��Ƥϡ����ڤ���Ƥ��ʤ��褦����
����ꥫ�⡢��ǯ11����졼��2�б����ڤ��ؤ����ƹ����ͽ�ɥ���(Centers for Disease Control and Prevention: CDC)�������줿�����ɥ����륹���Ȥˡ�Sanofi-Aventis�ʥ��Υե�-���٥�ƥ����˼Ҥ�������ꥫ���ܤȤη���θ��ˡ�H5N1���졼��2�ΥХ륯ǻ�̱դ���¤���Ƥ��롣
�ʤ�������Sanofi-Aventis�ʥ��Υե�-���٥�ƥ����˼һ����Υ��Υե����ѥ��ġ���Υ說����ϡ����졼�ɣ���°��������ȥʥ�������륹���˺�������Ƥ����ΤǤ��äƤ⡢���졼�ɣ���°���륦���륹���Ф��Ƥ⡢���Ϥ�����Ȥ��⤬����褦����
���ȡ�Bird flu vaccine may be more protective than thought��
�ɵ���2007/01/14�������ʤ�δĶ��ʤ�Ļ����ե륨����ĻĴ��
�Ķ��ʤȵܺ긩����Ļ����ե륨��ȯ����������Į���ܷܾ������������(2007ǯ1��16����ī��NHK�Ǥϡ�Ⱦ��300��ȥ����ƻ���줿�����Ķ��ʤ���˳�ǧ�����Ȥ�����Ⱦ��3������ȥ�Τۤ����������ȤΤ��ȤǤ��ä���)��Ĵ���ϰϤȤ��ơ���Ļ����©Ĵ����»ܤ���������Ļ�����̻�ʤɤΰ۾�ϸ�������ʤ��ä��Ȥ��Ƥ��롣
������������
Ĵ���оݤ���Ļ�ϡ���ǧ������Ļ������Τ������ޥ����ĥ��ߡ������Х�ʤɣ�������Ϥ�Ļ�ǡ���Ļ�μ������©���ʤɤ�Ĵ�٤���̡�����Ļ�˴ؤ��Ƥ����ʤξ������Ѥ��Ϥʤ��ä��פȤ�����
������������������ʤΤϡ���Ļ����ե륨��ȯ����������Į���ܷܾ�������������פ�Ĵ���ϰϤȤ����Ȥ������Ȥ���
�ȶ٤ϡ�Ⱦ��10������ȥ�����ư�ػߤʤΤˡ���Ļ��Ĵ���ϡ����ǡ�3��������(Ⱦ��3������ȥ�)�ʤ�?�äƤ��������Ѥʵ��䤬�襤�Ƥ��롣
����ˡ�����ξ��ˤϡ�9.3������ˡ���Ļ���罸����Ϥεܺ������������깾������Τˡ����������뤷�Ƥ��ʤ��褦�ʤΤǤ��롣
������Ͽޤϡ����Υ���������ɤ�����
�����������꤬��ư���¤��ϰϤǤ���10������������Ǥ��롣
�Ͽޤ��濴�ˡ�ë�������פȤ����Τ��������Ļ����ե륨��ȯ�����ǡ��Ͽޤα��Ф�夬���ܺ������Ļ�������ϤǤ����������깾�Ǥ��롣
��������깾�ξ���Ϥ����Ͽ��Ǥ��롣
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/08 Monday
2007/01/08
��
 ���������ܤγƿ�ʹ���������Ƥ����Ǿ���ɤ�ȯ��Ψ������ΰ����Ҥ������פȤ���ʸ�θ�ʸ�ϡ��������A nonsynonymous SNP in PRKCH (protein kinase C ) increases the risk of cerebral infarction��(Nature Genetics)�Ǥ��롣
���������ܤγƿ�ʹ���������Ƥ����Ǿ���ɤ�ȯ��Ψ������ΰ����Ҥ������פȤ���ʸ�θ�ʸ�ϡ��������A nonsynonymous SNP in PRKCH (protein kinase C ) increases the risk of cerebral infarction��(Nature Genetics)�Ǥ��롣
���פϡ����ΤȤ���Ǥ��롣
�ͤ��μ��θĿͺ��ϰ����Ź�Τ鷺���ʺ��ˤ�äƷ�ޤ뤬������ϡ����˥å�(�ӣΣ�-Single Nucleotide Polymorphism)�Ȥ������ΤǤ��롣
���˥åפϡ������Ҥα���������˰������Ѱۤ��������1%�ʾ�����٤Ǹ������Τ�����ɴ���������˰�Ĥγ�礤��¸�ߤ��롣
���줬���Ŀͺ������ݥ���ȤȤʤ��Τǡ����ܸ�Ǥϡ��ְ����¿���פȤ�����
����͵�彣�綵���������ظ����ε��ݽ��������������ϡ�52,608�ΰ����ҤΥ���SNP(�����ΰ��¿����¿����¸�ߤ��Ƥ����硢��������ɽ����ʬ�Ϥ��뤿������Ф줿SNP�Υ��å�)��ʬ�Ϥ�������ˡ����ܤ�ʡ������Ǿ���ɴ��Ԥ�ʡ������Į�η��Ԥ��줾�죱�������ͤ����ӡ���Ӹ������̡��٤�����ɤ��ͤޤ�֥饯��Ǿ����������( lacunar infarction )��ȯ�ɤ˴ؤ������Ҥ����������
���η�̡�Ǿ����(Ǿ������)�δ��ԤϷʿͤ���Ӥ��ơ��֥ץ��ƥ��ʡ����å�������(protein kinase C, eta)�ȸƤФ�뤿��Ѥ�����������Ҥ��������ʬ�������������֤�����äƤ���ͤ�¿�������ο͡��ϡ���������ΰ㤤��Ǿ����������ȯ��Ψ���⤯�ʤ뤳�Ȥ�ʬ���ä���
���ΣӣΣ�(�����¿��)�α����Фϥ��ǥ˥�ʣ��ˡ������˥�ʣǡˤΣ�����ǹ�������Ƥ��ꡢǾ���ɴ��ԤǤϷ��ʿͤ���١�������Ŀͤ���������¿���ä���
����ˡ���Į�ΰ��̽�̱�������ͤ�ǯ�֤ˤ錄�ä�����Ĵ�������Ȥ����������Ф��֣����פοͤϡ��֣ǣǡפοͤ��Ǿ����ȯ��Ψ���������ܹ⤫�ä���
���Ρ֣Уңˣãȡ�(protein kinase C, eta)�ȸƤФ��SNP(�����¿��)�ϡ�PKC��Protein kinase C�ˤγ�ư�˲��餫�αƶ���Ϳ���Ƥ����Τȸ����롣
�ޤ���PKC��Protein kinase C�ˤϡ���������˦�˸��졢�ʹ֤Υ��ƥ�����ư̮�Ų��ɾ㳲�ˤ����ơ�ˢ���Υޥ����ե������ȤʤäƸ��졢�㳲�Υ����פ���Ÿ����ˤĤ졢�����θ��ݤϡ����礷�Ƥ���Ȥ��Ƥ��롣
�Ȥ����ǡ����ä�������ʸ�˥������失��Ĥ��ϡ���Ƭ�ʤ��Τ�����������ǵ��ˤʤ�Τϡ�������͵�彣�綵���������ظ����ε��ݽ�����������������52,608�ΰ����ҤΥ���SNP��ʬ�Ϥ�������ˡ����ܤ�ʡ������Ǿ���ɴ��Ԥ�ʡ������Į�η��Ԥ��줾�죱�������ͤ����ӡ���Ӹ������̡פ���ʬ�Ǥ��롣
��ʸ�θ�ʸ�ˤ��С�
SNP����ϡ� the JSNP database���饻�쥯�Ȥ���
ʡ������Į�ξ���ϡ�14ǯ�֤ˤ錄��follow-up cohort study (��������ǯ����»ܤ��Ƥ������转���¤αֳ�Ĵ���ֵ�Į�����)��������줿�Ȥ��Ƥ��롣
�Ĥޤꡢ���θ���Υ١����ˤϡ������Ǿ���ɴ��Ԥȷ��Ԥΰ����Ҿ�����Ȥ������ȤǤ��롣
�����Ҹ�����������������Ҿ����Ŀ;����ݸ�ˡ�ˤ�äƼ����٤����פʸĿ;���Ǥ��ꡢ���Τ���ˤϡ��Ŀͤ����ꤹ�����Ȱ����Ҿ��Ĥʤ��뤳�Ȥ��ʤ��褦ƿ̾������ľ�ܸĿͤ�����Ǥ��ʤ��褦�ʻ��Ȥߤˤ��Ƥ��ʤ���Фʤ�ʤ���
�嵭�θ��������ʤΤϡ���ʡ������Į�η��ԡפȤ������Ȥǡ���̾�θ�ͭ̾���Ф��Ƥ��뤳�ȤǤ��롣
�Ĥޤꡢ����θĿͤȰ����Ҿ���Ȥ�����뤳�Ȥϡ����Ȥ��ɤ��Ƥ⡢�ϰ�Ȥ��ƤΥ��ۡ��ȥǡ����١���������ͭ����̾��ɽ�˽Ф��Ƥ��뤳�Ȥˤ�äơ�����Ū�ʥ������Ƥ��뤳�ȤǤ��롣
�⤷�����줬�������������Ū�и��ˤĤʤ��뤳�Ȥ����äƤϤ����ʤ��������Τ褦�ʸ���ξ��ˤϡ��������̾�����餫�ˤ��뤳�Ȥϡ����ۤ��������ΤǤϤʤ���������
�ʤ������ͤޤǤˡ����Υ����ȡ�ʿ��16ǯ�ٲʳص��ѿ���Ĵ������ƳŪ�������ο���ֵ�Į�ˤ��������转���¤Υ��Υ�ֳظ���»ܷײ��פˤϡ����Τ褦�˽�Ƥ��롣
��40�аʾ�ε�Į��̱����¿�����оݤ����٤ι⤤��Ǥ�Ԥ�����ե���ץ��3�ʹ�Ʊ�������ؿˤˤ������äƼ������������ε�Į����Υǡ����ȤȤ�ˡ����ο����ʽ��Ĥ�������ǡ�˶������Ӱ���������Į�إ륹C��C�����ˤ����ƥǡ����١���������ȤȤ�ˡ�Ʊ������˥ǡ����١����δ��������ƥ���ۤ��롣��
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/05 Friday
2007/01/05(Fri)
��
 ����ī����ʹ�Υ��������Ҳ����������褦�פϡ��䤬���������פäƤ��뤳�Ȥ����ۤ��Ƥ��롣
����ī����ʹ�Υ��������Ҳ����������褦�פϡ��䤬���������פäƤ��뤳�Ȥ����ۤ��Ƥ��롣
�Ĥޤꡢ��졢�֤����βǤˤ⡢���Ҳ��к����פʤɤȡ����ν������Ҥ���Ū������ɤ��������ˡ��ष�������Ҳ��ˤʤ�Ⱥ����������������ľ�����Ȥ���Ϥ�ʤ���Фʤ�ʤ��ΤǤϤʤ����������Ȥ�������ռ��Ǥ��롣
���Ҳ��ˤʤ�ȡ�����������������Ȥϡ����������������夬��ǡ�����ޤǡ�����Ω�äƤ�����������Ȥ������ȤǤ��롣
����äȡ����Ȥ��ϰ��������ޥ����ˡ�ǡ���̳��ؤβ��(���åץ饤��)�ˡ����Ū�˹פ����Ȥˤʤ벼�̳��ؤβ���ʥ�����饤��ˤ������ʤ��褦�ʾ��֤Τ�Τȡ��ͤ�����ä��ᤤ��������
ǯ��ϡ��ޤ��ˤ������������ϤηкѤ�����Ω�äƤ��륹������ϡ�¿���쾯�ʤ��졢���αƶ����롣
�����кѤ������ϡ����⤽�⡢���²��ǵ��ϤηкѤ�����Ω���ʤ��кѷ��ˡ����Ϥηкѥ��åȤ�����Ω�äƤ�����ꥹ���ѡ������������Ƥ������Ȥˤ�롢���饦�ǥ���������Ū�����Ǥ⤢�롣
�ޤ�������ޤǤΡ�����ե����פ��ƤΡ�������μ¼���ô�ڸ��Υ����������þ������ˤ��ƤΡ�����������������Ȥ�����Τͤ��Ƥ���ɬ�פ�����ΤǤϤʤ���������
����ϡ��°פʡ��ǥե�æ�����䡢����Ū�ʥ���ե쥿�����å��������æ�ѤǤ��ꡢ��������ʡ�����ե�ָ�����β���Ǥ⤢�롣
���⤽�⡢ǯ��Υ��������ޤޤʤ�ʤ��Τˡ������Ǥ�����ǯ������Ԥ�������ե��������ɸ�Ȥ��롢��Υ��������������ե쥿�����å����ˤϡ����롣
���Τ褦�ʴ������顢���Ҳ��ˤ��ä����Ƽ���������������Ԥ߽Ф��Ƥ������Ȥ��������줫��ɬ�פʤ��Ȥ����Τ�ʤ���
���줾����å����ե�����(��å����ե�����)�ؤξ��Ҳ��к������뤤�ϡ������饻��ؤξ��Ҳ��к��ؤΡ��ѥ�����ॷ�եȤȤ����٤���?
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
2007/01/02 Tuesday
2007/01/01(Mon)
��
 �ּ�ʬ�Τ��������Ф��Τˡ����������Ҥ���¿���������ʧ��ʤ���Фʤ�ʤ��ΤϤ���������
�ּ�ʬ�Τ��������Ф��Τˡ����������Ҥ���¿���������ʧ��ʤ���Фʤ�ʤ��ΤϤ���������
����ϡ������������ԤΥ��åɡ��Хå�Ƭ��θ��դǤ��롣
���ȡ���ԣ��ԣּ͡��ס��������פ�̵����������
���ߤ����⡢Ŭ�ʤʸ��դ�Ҥ٤���Τȡ��������롣
�����������Ԥϡ�ATM����������Ρ֥����Хץ����ӥ���Ϥ���Τ�����������Ф��ơ���ɩ���UFJ��ԡ�MUFG�ˤʤɤ��֤�����μ������ô�������פȤ��ߤĤ�������ߤ��ʤ��ʤ顢ATM��Ȥ�������פȤ������������������Ԥϥ����Х����ӥ����³���Ƥ��롣
����ʡ����ܤζ�ͻ���ξ��Ⱥ����˸����줿�и���Ƨ�ޤ��ƤΥ��åɡ��Хå�Ƭ�����Ƭ�Τ褦�ʸ��դʤΤ�������
�̶�ԤΤ������ˤĤ��Ƥϡ����Τۤ��ˤ⡢���ͤƤ��餳�Υ֥����Ǥ�����Ȥ��Ƥ��뷫�����Ƕ�����꤬���롣
��Ԥ����ɺĸ��ν����侭���ȯ���������ư�������Ѥ�Τ��������ΤȤ��ˡ��Ƕ��Ǽ��롣
ͻ����ֺѤ��ʤ���»�������������ˤϡ���Ԥ���˰������Ƥ�������Ƿ���᤹��Τ��������κݡ��������������Ѥ������ΤȤ���Ʊ����Ǽ��Ǥ���äƤ��롣
�ᤷ���ϡ���Ԥ������ˤʤä������ǡ�����ǯ��Ǽ�dzۤ��麹��������뤳�ȤǷ�Ѥ���롣
����ϡ�������äƤ����Ƕ�ˤĤ����ղû���Ǥ��롣
����ˤĤ��Ƥϡ�2002ǯ10��31���Υ֥�������Ԥ��Ǹ��̲�פǡ���Ψ����?�δ��ղû��⤬�����äƤ���?�פǽҤ٤��褦�ˡ��������+����(�����������ǯ7.3��)�Ȥ��������汿�Ѥ��⡢���̤˹⤤���Ҥ��Ĥ��Ƥ����äƤ��뤳�ȤǤ��롣
�֤��뤫�֤��ʤ�������̯�ʥ��졼������ĸ��������ߤ��Ϥ����������ɺĸ��˻�Ω�ƾ夲�������ӥ����˥���ĸ��Ȥ��ơ��¤��Хå���ꤷ�ơ�������Ф���������Ф��ۤɡ�������Ǽ��Ƕ�˹�������©���Ĥ��ơ����ղû���Ȥ��ơ����äƤ��롢�Ȥ������ޤ���
�ʤ�Ȥ⡢���ԿԤ��äǤϤ���ޤ���?
为��译对汉语, ����这
��http://translate.livedoor.com/chinese/
Translate
��http://www.google.com/translate_t
��������HOME-���ԥ˥���-���-����-ȯ��-�ץ��ե�����-�Ǽ���-���ո�
{kind=link}
{kind=link}
{kind=link}
{kind=link}